146 / BIOLOGÍA DEL SISTEMA INMUNITARIO
El sistema inmunitario es una red de componentes celulares y solubles que interaccionan entre sí. Su función es distinguir entidades dentro del cuerpo como «propias» o «extrañas» y eliminar las extrañas. Los microorganismos son las principales entidades extrañas, pero también son importantes las neoplasias, los trasplantes y ciertas sustancias extrañas (p. ej., algunas toxinas). Para desarrollar esta tarea, el sistema inmunitario ha desarrollado dos mecanismos: la inmunidad inespecífica y la inmunidad específica, que están ligadas entre sí e influyen la una en la otra.
Inmunidad inespecífica (innata)
Este tipo de inmunidad es la más antigua desde el punto de vista filogenético, está presente desde el nacimiento, no requiere un encuentro previo con la sustancia ofensiva y no da lugar a memoria. La inmunidad innata comprende las barreras, como la piel, y la protección química, como el ácido gástrico. Existen dos componentes celulares: 1) el sistema fagocítico, cuya función es ingerir y digerir los microorganismos invasores, y 2) las células agresoras naturales (NK, del inglés natural killer), cuya función es destruir ciertos tumores, microorganismos y células infectadas por virus (v. más adelante). Los componentes solubles constan de proteínas del complemento, reactantes de fase aguda y citocinas.
Los fagocitos son los neutrófilos, los monocitos (en la sangre) y los macrófagos (en los tejidos). Los macrófagos, que están ampliamente distribuidos, se sitúan de forma estratégica en las interfases de los tejidos con la sangre o las cavidades; por ejemplo, los macrófagos alveolares (pulmones), las células de Kupffer (sinusoides hepáticos), las células sinoviales (cavidades articulares), las células de la microglía perivascular (recubrimiento del SNC) y los fagocitos mesangiales (riñones).
Las citocinas son polipéptidos diferentes a las inmunoglobulinas que secretan los monocitos y los linfocitos en respuesta a la interacción con un antígeno específico (Ag), un Ag inespecífico o un estímulo soluble inespecífico (p. ej., endotoxina, otras citocinas). Las citocinas afectan a la magnitud de las respuestas inflamatorias o inmunitarias. Aunque la secreción de citocinas puede activarse por la interacción de un linfocito con su Ag específico, las citocinas no son específicas para el antígeno; de este modo, unen las inmunidades innata y adaptativa.
Inmunidad específica (adaptativa)
La inmunidad específica se caracteriza por el aprendizaje, la adaptabilidad y la memoria. El componente celular es el linfocito y el componente soluble las inmunoglobulinas (Ig).
Los linfocitos se dividen en dos subgrupos: los derivados del timo (células T) y los derivados de la médula ósea (células B). Los linfocitos tienen una distribución clonal; cada clon se especializa en el reconocimiento de un Ag específico mediante su receptor para el Ag. Debido a que el número de Ag no tiene en teoría límite, esta especialización podría representar una sobrecarga excesiva para el sistema inmunitario. El dilema de proporcionar un número infinito de clones únicos se resuelve mediante la capacidad de los genes del receptor para el Ag del linfocito de combinarse de un número de formas en teoría ilimitado.
La función de receptor del Ag en las células B está mediada por las inmunoglobulinas de superficie (Igs). Después de que las células B se unen al Ag soluble a través de sus Igs, una serie de acontecimientos (p. ej., proliferación, diferenciación) culminan en la secreción de Ig, que es el anticuerpo específico (Ac) para ese Ag. La idea actual es que el repertorio de Ac de un organismo antes de la exposición al Ag se debe a los Ac generados durante la maduración de la células B por los reordenamientos genéticos de las Ig. Para comprender la naturaleza del ordenamiento genético de las Ig es necesario conocer su estructura (v. también Estructura del anticuerpo, más adelante).
Las Ig se componen de dos cadenas pesadas y dos ligeras, cada una con dominios constantes y variables. El Ag se une al dominio variable. A nivel genético, la región C está codificada por los genes de la región C; la región V (para las cadenas ligeras) por los genes de las regiones V y J y (para las cadenas pesadas) por los genes de las regiones V, D y J. Estos segmentos genéticos no se sitúan a continuación uno del otro en el cromosoma, sino que están separados y deben yuxtaponerse durante la maduración de la células B. De este modo, para sintetizar una cadena pesada, uno de los varios segmentos D (se han identificado al menos 12) se une a uno de los seis segmentos J. Este grupo se une a su vez a uno de los varios cientos (posiblemente miles) de segmentos genéticos de la región V para formar una unidad transcripcional completa para una cadena pesada de Ig.
Dependiendo de qué segmento de cada región genética se utilice, es posible crear un número enorme de moléculas de Ig con diferentes especificidades. La posible diversidad aumenta aún más por la adición de nucleótidos aleatorios en los lugares de unión (entre las regiones V, D y J), mediante mutaciones somáticas puntuales y por las imprecisiones en la unión de los diferentes segmentos. Se cree que el repertorio de Ac de un organismo antes de exponerse al Ag se debe a los Ac generados antes de la maduración de la células B, durante los reordenamientos genéticos de Ig.
Las células T no tienen Igs pero reconocen el Ag mediante su principal herramienta de reconocimiento, el receptor de la célula T (TCR, del inglés T-cell receptor), y otras moléculas de adhesión accesorias. Los genes que codifican el TCR pertenecen a la superfamilia de los genes de Ig; de forma similar a los genes de Ig, se recombinan dando lugar así a un gran número de clones de células T, cada uno con una respuesta a Ag específica.
La porción que se une al Ag del TCR consta de dos cadenas (ab o gd); cada una tiene un dominio constante y un dominio variable. Al contrario que la IgE, que existe de forma independiente sobre la superficie de la células B, el TCR se asocia con la molécula CD3; a la unidad completa se la denomina complejo TCR/CD3. Aunque las cadenas TCR está sujetas a un reordenamiento genético y son variables, las cadenas CD3 (que constan al menos de cinco cadenas) son invariables y no son específicas para el antígeno. Algunos Ac anti-CD3 activan las células T directamente, eludiendo así la necesidad del Ag. Por ello, el CD3 es importante para la transducción de la señal de activación a través de la membrana del linfocito.
Los linfocitos pueden dividirse a su vez en subgrupos bien por función o bien por los marcadores de superficie. Los subgrupos de linfocitos se han identificado mediante combinaciones de ciertas moléculas de su superficie. Estos marcadores de superficie se han denominado grupos de diferenciación (CD, del inglés clusters of differentiation). Hasta la fecha, se han identificado 166 CD. En Internet puede encontrarse información actualizada sobre los antígenos CD (http://www.ncbi.nlm.nih.gov/prow).
Complejo principal de histocompatibilidad
La capacidad del sistema inmunitario de diferenciar lo propio de los extraño está determinada en gran medida por los productos del complejo principal de histocompatibilidad (MHC, del inglés major histocompatibility complex) cuyos genes están situados en el cromosoma 6, pertenecen a la superfamilia de genes de Ig y pueden recombinarse. Los productos del MHC de la clase I constan del HLA-A, B y C; tienen una distribución amplia y están presentes en la superficie de todas las células nucleadas y de las plaquetas. Los productos del MHC de la clase II constan del HLA-D, DR, DP y DQ; tienen una distribución más limitada en las células B, los macrófagos, las células dendríticas, las células de Langerhans y las células T activadas (pero no en reposo).
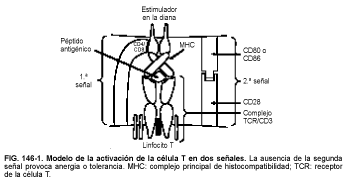
Las células B pueden responder a Ag solubles, pero las células T raramente lo hacen y reconocen al Ag sólo cuando está incluido dentro del MHC; por tanto, las células T reconocen complejos MHC/Ag. El mecanismo por el cual se procesa el Ag y se asocia con el MHC antes de presentarse a las células T tiene lugar en las células presentadoras de antígeno (APC, del inglés antigen presenting cells), por ejemplo, las células de Langerhans, los monocitos, los macrófagos, las células dendríticas foliculares y las células B. Aunque los matices no se conocen por completo, parece que es necesario que el Ag esté desplegado, degradado y fragmentado para ser procesado. Mediante un proceso exógeno, el Ag sufre endocitosis y degradación en los lisosomas, se asocia con productos del MHC de la clase II y se transporta a la superficie celular. Mediante un proceso endógeno, el Ag se produce dentro de la célula (p. ej., por una infección viral) y sufre una degradación fuera de los lisosomas en organelas llamadas proteasomas. Los péptidos resultantes se transportan al retículo endoplásmico rugoso (RER) mediante proteínas transportadoras. Una vez en el RER, estos péptidos se asocian con los productos del MHC de la clase I antes de ser transportados a la superficie celular. Es importante saber si el Ag se asocia al MHC de la clase I o II, porque las moléculas CD4 y CD8 actúan como moléculas de adhesión accesorias al unirse a las clases II o I, respectivamente. La unión del TCR con el complejo MHC/Ag puede no ser suficiente para inducir la activación de las células T. Es necesaria una señal de coactivación; esta segunda señal está mediada por la unión del CD28 de la superficie de la células T con CD80 o CD86 de la APC. La ausencia de una interacción CD28/CD80-CD86 puede hacer a la célula T anérgica o tolerante (v. fig. 146-1)
Citocinas
Aunque es necesario un contacto celular íntimo para que las células T respondan de forma óptima, éstas y los monocitos secretan citocinas que pueden influir en acontecimentos cercanos o distantes. Las citocinas interaccionan con receptores de superficie celular específicos y pueden actuar de una forma autocrina o paracrina.
Las citocinas pueden dividirse en varios grupos, entre los que se encuentran los interferones (IFN-a, b y g), el factor de necrosis tumoral (TNF-a y b), las interleucinas (IL-1 a IL-8), los factores transformadores del crecimiento y los factores estimuladores de colonias (CSF, del inglés colony stimulating factor). Ver en la tabla146-1 las principales citocinas, sus principales orígenes y sus principales efectos.




Aunque las diferentes citocinas y sus efectos suelen enumerarse por separado, es importante recordar que las citocinas actúan en conjunto, en tándem o en conflicto con una respuesta inmunitaria dada; por ejemplo, la IL-1 puede inducir la secreción de IL-2. La IL-2, la IL-4 y la IL-6 pueden actuar de forma sinérgica generando linfocitos T citotóxicos. La IL-4 y el IFN-g pueden actuar contrarrestándose mutuamente en la inducción de la expresión de la clase II en las células B y en la inducción de la secreción de IgE.
La orquestación simultánea de varias respuestas y la redundancia del sistema inmunitario queda quizá mejor ilustrada a través de la estructura de algunos de los receptores de interleucinas. El receptor de la IL-2 consta de tres cadenas: a, b y g. La expresión de las tres cadenas da lugar al receptor de IL-2 de alta afinidad; la expresión de las cadenas b y g da lugar sólo a un receptor de IL-2 de afinidad intermedia, mientras que la cadena a representa sólo un receptor de afinidad baja. Recientemente se ha demostrado que las mutaciones o eliminaciones de la cadena g del receptor de la IL-2 constituyen la base molecular de la inmunodeficiencia combina grave ligada al X (IDCG). Resulta interesante señalar que las mutaciones de las cadenas a o b del receptor de la IL-2 no dan lugar a la IDCG (al menos en modelos animales). Esta aparente discrepancia se debe a que la cadena g del receptor de la IL-2 es también parte del complejo receptor para la IL-4, la IL-7, la IL-9 y la IL-15; esta cadena se denomina ahora cadena g común (gc). El receptor de la IL-15 comparte las cadenas b y g con el receptor de la IL-2. La cadena a del receptor de la IL-13 es idéntica a la cadena a del receptor de la IL-4. Los receptores de la IL-3, la IL-5 y el GM-CSF tienen todos una cadena b idéntica.
Una nueva familia de citocinas es la denominada de forma acertada quimiocinas; las quimiocinas inducen la quimiotaxis y migración de subgrupos de leucocitos. Hay cuatro subgrupos de quimiocinas, que se definen por el número de aminoácidos intermedios ente los primeros dos residuos cisteína de la molécula. Algunos de los receptores de las quimiocinas pueden servir de correceptores para la entrada del VIH en los monocitos/macrófagos.
CÉLULAS T E INMUNIDAD CELULAR
Las células T maduran, adquieren repertorios funcionales y aprenden el concepto de lo propio en el timo. El timo desarrolla las dos tareas de selección positiva (a los clones que reconocen el Ag/MHC se les permite proliferar, madurar y migrar a la periferia) y selección negativa (se eliminan los clones que reaccionan a lo propio como si fuera extraño). No se conocen por completo los mecanismos celulares y moleculares exactos de esta selección.
Durante el desarrollo fetal, la célula T primordial (stem cell), que deriva de la médula ósea, se traslada al timo, donde madura y aprende el concepto de lo propio. Tiene lugar el proceso de la selección tímica y a los linfocitos maduros se les permite dejar el timo; se les encuentra en la sangre periférica y en los tejidos linfoides. Todas las células T maduras expresan CD4 o CD8 de una forma mutuamente exclusiva.
Células T cooperadoras
A las células T que expresan el CD4 se les denomina generalmente linfocitos T cooperadores (TH). Estas células pueden subdividirse en dos categorías principales, dependiendo de su función, respuesta a diferentes citocinas y capacidad para secretar citocinas. La opinión actual es que las células TH comienzan como células precursoras que sintetizan IL-2. Ante una estimulación inicial, estas células evolucionan a células THO, que pueden secretar varias citocinas, como IFN-g, IL-2, IL-4, IL-5 y IL-10. Dependiendo de la citocina disponible, las células THO pueden evolucionar a células TH1 o TH2; el IFN-g y la IL-2 favorecen el desarrollo a TH1 y la IL-4 y la IL-10 favorecen el desarrollo a TH2. Las TH1 y las TH2 difieren en el perfil de citocinas que secretan: las células TH1 secretan IFN-g, mientras que las células TH2 secretan IL-4, aunque ambas secretan otras citocinas (p. ej., IL-3, GM-CSF, TNF-a). En general las TH1 favorecen la inmunidad celular, mientras que las TH2 favorecen la inmunidad humoral.
La distinción de las respuestas TH1 y TH2 ha cambiado la idea acerca de la relación del sistema inmunitario con la enfermedad. La respuesta inmunitaria debe ser no sólo fuerte, sino también adecuada para la infección o la enfermedad. Quizá el mejor ejemplo de esta estrategia es la lepra, en la cual ahora se cree que la respuesta TH1 da lugar a la lepra tuberculoide, mientras que la respuesta TH2 origina la lepra lepromatosa. Además, una respuesta TH1 puede agravar las enfermedades autoinmunitarias, mientras que una respuesta TH2 favorece la secreción de IgE y el desarrollo de atopia.
Células T supresoras/citotóxicas
Las células T que expresan CD8 están peor caracterizadas que los subgrupos TH, aunque parece que también pueden dividirse en dos tipos dependiendo de las citocinas que secretan, siendo la segregación idéntica a la de los subgrupos CD4. Se ha sugerido que a los tipos de linfocitos se les llame tipos 1 y 2 (T1, T2) en lugar de TH1 y TH2, porque puede hacerse la misma subdivisión en las células CD8.
Las células T citotóxicas (TC) se refieren a linfocitos T citotóxicos restringidos por el MHC y específicos frente al Ag (CTL, del inglés citotoxic T lymphocytes; v. más adelante). Las células CD4 y CD8 pueden actuar como CTL dependiendo de si reconocen a las clases I o II del MHC, respectivamente. También se reconocen varios tipos de células citotóxicas o agresoras; algunas de ellas expresan marcadores CD8 o CD4.
Células agresoras
La identificación de cada tipo (o de varios) depende de la restricción por MHC, la necesidad de sensibilización, la especificidad de la diana y las respuestas a las citocinas. Aunque los macrófagos pueden ser citotóxicos, esta toxicidad es inespecífica y se debe a la activación de algunas citocinas. Los diferentes tipos de células agresoras pueden dividirse de una forma sencilla en las restringidas por el MHC (p. ej., CTL) y las no restringidas por el MHC (p. ej., células NK). Ningún tipo necesita Ac, complemento ni fagocitosis para matar a la célula diana. En su lugar, producen una señal lítica a través de la célula diana tras establecer un contacto íntimo de célula a célula.
Células agresoras restringidas por el MHC. Los linfocitos T citotóxicos (CTL) son células agresoras que se generan sólo en la sensibilización específica, bien contra células que expresan productos del MHC extraños (alogénicas) o bien frente a células autólogas, siempre que estas células hayan sido modificadas por una infección viral o un hapteno químico (singénicas). La vida de un CTL tiene tres fases: una célula precursora puede convertirse en citotóxica con una estimulación apropiada; una célula efectora se ha diferenciado y puede lisar su diana adecuada, y una célula de memoria, en reposo y sin estimulación, está preparada para convertirse en efectora tras una nueva estimulación por las células originales. Las células intactas son los estimuladores más potentes para la generación de CTL; el Ag soluble es ineficaz excepto bajo ciertas condiciones. Como se dijo antes, el Ag se procesa y fragmenta y se introduce en la hendidura presentadora del Ag del MHC. Ahora es posible identificar los péptidos que se ajustan perfectamente a diferentes haplotipos del MHC. Si se utilizan tales péptidos para la estimulación, pueden incluirse en el MHC y de este modo estimular una respuesta de células T.
Los CTL alogénicos pueden generarse fácilmente in vitro en cultivos de linfocitos normales en células estimuladoras alogénicas irradiadas que difieren en todo o en parte en el MHC. Los CTL alogénicos pueden también generarse in vivo en trasplantes de un órgano derivado de un donante cuyos productos del MHC difieren de los del receptor, lo que probablemente desempeñe un papel importante en el rechazo de los órganos trasplantados. La generación satisfactoria de CTL requiere dos señales: la señal antigénica (células estimuladoras) y la señal de amplificación (citocinas). Una acción eficaz de estas dos señales requiere APC, TH y precursores de TC. La señal de amplificación está mediada por citocinas que actúan en tándem; las más importante son la IL-1, la IL-2 y la IL-4. Se cree que en la generación de CTL intervienen otras citocinas (incluidas la IL-6, la IL-7, la IL-10 y la IL-12), al menos in vitro.
Otro tipo de CTL que es importante para eliminar ciertos agentes patógenos intracelulares (especialmente en células infectadas por virus) es el también llamado CTL específico para el antígeno (CTL singénico). El CTL singénico reconoce sólo células diana que expresan el antígeno utilizado para la estimulación asociado con el MHC. Estos CTL se generan frente a células autólogas siempre que las células se hayan «modificado» por una infección viral o haptenos químicos. La expresión de productos virales, o haptenos, en la superficie celular asociados con el MHC desencadena una cascada de diferenciación celular y liberación de citocinas y una respuesta similar al CTL alogénico. Los CTL alogénicos y singénicos utilizan el complejo TCR/CD3 para reconocer a la célula diana.
Células agresoras no restringidas por el MHC. Al contrario que las CTL, las células agresoras naturales (NK, del inglés natural killer) no necesitan estar sensibilizadas para expresar su función agresora. Las células NK constituyen el 5-30% de los linfocitos de la sangre periférica. Las células NK son linfocitos, pero no pertenecen a las líneas de células T o B. Por tanto, las células NK no expresan Ig ni TCR/CD3 en su superficie. Los marcadores de superficie que mejor caracterizan a las células NK son CD2+, CD3-, CD4- y CD56+, y un subgrupo es CD8+. Las células NK matarán a células autólogas, alogénicas e incluso a células tumorales xenogénicas sin importar si expresan o no el MHC; de hecho, pueden matar de forma preferente células diana que expresen poco o ningún tipo de la clase I del MHC. La susceptibilidad al efecto agresor de las células NK puede reducirse si se induce a la célula diana a aumentar la expresión de su MHC (p. ej., mediante transfección o con IFN).
Esta aparente inhibición de la actividad agresora NK por la expresión de la clase I del MHC condujo a identificar varias clases de receptores de la clase I del MHC en la superficie de las células NK. Estos receptores tienen una estructura diferente a la del TCR y en general se les denomina receptores inhibidores de la célula agresora (KIR, del inglés killer inhibitory receptors). Mientras que la unión del MHC con el TCR de las células T activa a la célula T, la unión del MHC con la mayor parte de los KIR inhibe la actividad NK, aunque algunos KIR pueden producir activación. También se han identificado los KIR en las células T. Esto representa un enigma interesante: las células T tienen receptores diferentes (TCR/CD3 y KIR) para la misma molécula (clase I del MHC) pero con efectos opuestos. No se conoce con exactitud qué decide la activación o inhibición de la célula T, y el resultado puede variar dependiendo del clon de células T.
Durante mucho tiempo se ha pensado que las células NK eran importantes en la vigilancia tumoral porque pueden matar a algunas células diana tumorales y porque la mayor parte de los tumores no expresan el MHC. Las células NK también matan a algunas células infectadas por virus y a algunas bacterias (p. ej., Salmonella typhi). Sigue sin aclarase la estructura del reconocimiento del Ag de las células NK.
Además de su propiedad agresora, las células NK pueden secretar varias citocinas, sobre todo IFN-g y GM-CSF (factor estimulante de colonias de granulocito-macrófago). Las células NK pueden ser la fuente más potente de IFN-g. Al secretarlo, las células NK pueden influir en el sistema inmunitario adaptativo al favorecer la adaptación de los TH1 e inhibir la diferenciación de los TH 2.
Citotoxicidad dependiente de anticuerpos
Las células NK expresan el CD16, un receptor para el Fc de la IgG (v. Estructura de anticuerpos, más adelante) y pueden utilizar este receptor para mediar otro tipo de lisis celular no restringida por el MHC. La citotoxicidad celular dependiente de Ac (ADCC, del inglés Ab-dependent cell-mediated cytotoxicity) depende del Ac que reconozca una célula diana (por tanto, la especificidad de la ADCC viene dada por la especificidad del Ac). Tras unirse a su Ag, la región Fc del Ac se expone y se unirá a su receptor en la célula NK para formar un puente. Una vez que se forma el puente se produce una señal lítica poco conocida en la célula diana, que provoca su muerte.
Una forma interesante de ADCC es la también denominada ADCC inversa. Ciertas células agresoras, incluidos los CTL restringidos por el MHC, que expresan CD3 en su superficie, pueden perder la especificidad en presencia de Ac anti-CD3. El anti-CD3 se une a su ligando en la superficie de la célula agresora, dejando su porción Fc libre para unirse a las células diana que portan receptores del Fc. De nuevo, una vez que se forma el puente, se produce la señal lítica a la célula diana que lleva el Fc. Algunas formas de ADCC podrían ser útiles para marcar algunas células tumorales in vivo como forma de inmunoterapia.
Células T agresoras no restringidas por el MHC
Además de las células NK que son CD3- TCR- CD56+, otro subgrupo es CD3+ CD56+ que puede expresar CD2, CD5 y CD8. La mayoría tienen el TCR-gd, aunque se han identificado algunos clones TCR-ab. Este subgrupo puede mediar alguna actividad espontánea del tipo NK y puede aumentar este tipo de actividad tras la estimulación con IL-2. Otro subgrupo de células T (CD3+ TCR-gd CD4- CD8- CD56- CD16-) puede ser citotóxico, aunque la mayoría son clones o líneas celulares. Seguimos sin saber si los leucocitos frescos aislados con este fenotipo muestran actividad citotóxica espontánea.
Células agresoras activadas por linfocinas
Algunos linfocitos cultivados con IL-2 evolucionan a potentes células agresoras activadas por linfocinas (LAK, del inglés lymphokine-activated killers) capaces de lisar una amplia variedad de células diana tumorales, así como linfocitos autólogos que han sido modificados mediante cultivo, ciertos virus o haptenos. Las LAK se contemplan como un fenómeno en lugar de como un subgrupo de linfocitos bien definido. Los precursores de las LAK son heterogéneos, pero pueden dividirse en dos categorías principales: similares a NK y similares a T. En general están de acuerdo en que las células NK clásicas son los principales precursores de las LAK en la sangre periférica, pero esto puede no ser cierto en los tejidos extravasculares.
Pruebas de la inmunidad celular
Una evaluación cuantitativa mínima de la inmunidad celular debe incluir un recuento de linfocitos, el número de subgrupos de células T (CD3, CD4, CD8) y el número de células NK por análisis fluorescente. La evaluación cualitativa consta de las pruebas cutáneas de hipersensibilidad retardada (HR) y la siguientes pruebas in vitro: 1) proliferación en respuesta al Ag soluble, al Ac anti-CD3 y a alo-Ag; 2) la actividad lítica de las células NK espontánea y tras estimulación con IL-2 o IFN; 3) la capacidad de elaborar citocinas poniendo énfasis en el IFN-g, el TNF-a, la IL-2 y la IL-4, y 4) la capacidad para generar CTL restringidos por el MHC. Un análisis más profundo dependerá de los resultados de estas pruebas. Las pruebas completas de la inmunidad celular se limitan a los laboratorios de investigación.
Las pruebas cutáneas de HR establecen la normalidad de algunos aspectos del sistema inmunitario celular. Sin embargo, no estudian el estado de las células CD8, las células CD4 vírgenes, las células NK ni las células APC que no sean células de Langerhans. Por ejemplo, un paciente puede tener una ausencia completa de células NK y presentar todavía una HR normal. De este modo, aunque unas pruebas cutáneas de HR negativas indican una inmunidad celular anormal, lo inverso no es cierto (v. Redes inmunitarias, más adelante).
Las pruebas cutáneas de HR deben leerse a las 48 h. Una respuesta más precoz podría deberse a una reacción de Arthus (que comienza de 4-6 h después de la prueba y puede estar presente hasta 24 h). Una reacción de Arthus se debe a la presencia de Ac que se unen al Ag inyectado, lo que da lugar a la formación de un complejo inmunitario, la activación del complemento y la quimiotaxis de neutrófilos. El infiltrado celular de una reacción de Arthus consta sobre todo de neutrófilos, mientras que el infiltrado en la HR se compone de células mononucleares. La respuesta de HR comienza a resolverse pasadas 48 h, y si se lee la prueba cutánea a las 72 h, una reacción positiva en el límite (>5 mm de induración) puede parecer negativa.
REDES INMUNITARIAS
El sistema inmunitario opera como un todo, ningún componente actúa de forma autónoma. En cualquier respuesta inmunitaria los componentes actúan en conjunto, en tándem o en conflicto, como muestra la capacidad del sistema inmunitario de eliminar microorganismos. Los microorganismos extracelulares (la mayor parte de las bacterias encapsuladas) necesitan sólo ser fagocitadas para ser digeridas; sin embargo, los microorganismos intracelulares (p. ej., las micobacterias) son fáciles de ingerir, pero no pueden ser digeridas a no ser que el macrófago reciba una señal de activación.
La estrategia de eliminar los microorganismos extracelulares se dirige, por tanto, hacia la fagocitosis, que está facilitada por la opsonización (cubierta de un microorganismo con Ac, productos del complemento o ambos). Debido a que la mayor parte de los fagocitos tienen receptores para la fracción Fc del Ac y para los productos C3, la presencia de estas moléculas sobre una bacteria facilita su adhesión e ingestión. Esta respuesta inmunitaria «simple» depende de una síntesis satisfactoria de Ac, de la activación de la cascada del complemento y de un sistema fagocítico intacto. Los Ac los sintetizan las células B, aunque éstas están sujetas a la cooperación y supresión recibida por las células T. Además, se reclutan fagocitos mediante factores quimiotácticos, algunos de los cuales son sintetizados por las células T.
La estrategia de eliminar algunos microorganismos intracelulares que infectan a los fagocitos implica la activación de células del huésped, que después «se arman» y son capaces de lisar a estos microorganismos de una forma inespecífica. La capacidad de activar los macrófagos se encuentra en el corazón de la reacción de hipersensibilidad retardada (HR) típica, y la prueba cutánea de HR es un ejemplo excelente de las diferentes cascadas en una respuesta inmunitaria dada. La premisa de una prueba cutánea de HR es que la inyección intradérmica de un Ag al cual el paciente se ha expuesto previamente provoca una induración local en menos de 48 h. La intrincada red implicada en tal respuesta se ilustra en la figura 146-2. Tras la inyección, las células de Langerhans de la piel captan el Ag, lo procesan y lo presentan (formando un complejo con la clase II del MHC) a una célula CD4+ que se expuso previamente al Ag (es decir, una célula memoria de vida larga). Una vez que la célula CD4+ se une al complejo Ag/MHC, expresa los receptores de la IL-2 y libera varias citocinas (p. ej., IFN-g, IL-2 y factores quimiotácticos de linfocitos y macrófagos). El IFN-g induce a las células endoteliales a aumentar su expresión de células de adhesión, facilitando así la salida de linfocitos y macrófagos a través de la barrera endotelial. La IL-2 y la IFN-g también actúan como señales de proliferación/diferenciación, permitiendo expandirse a los clones memoria de las células T y a las células T que acaban de llegar. Una vez que los macrófagos alcanzan el lugar de la inyección, se evita que dejen esta zona mediante los factores inhibidores de la migración (MIF, del inglés migration-inhibiting factors) secretados por las células T activadas. El IFN-g y el GM-CSF, ambos secretados por las células T, actúan entonces como factores activadores de macrófagos (MAF, del inglés macrophage-activating factors). Los macrófagos activados están ahora «armados» y pueden lisar microorganismos intracelulares y cualquier célula tumoral próxima.
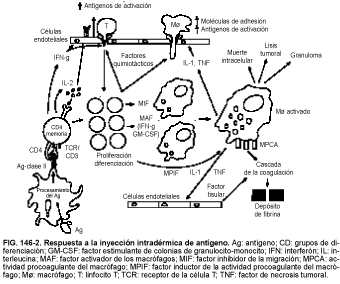
Los macrófagos activados secretan IL-1 y TNF-a que potencian la secreción de IFN-g y GM-CSF, aumentan la expresión de las moléculas de adhesión en las células endoteliales y permiten a estas células secretar factor tisular, que activa la cascada de la coagulación, que termina con el depósito de fibrina. De forma simultánea, los linfocitos activados secretan factor inductor procoagulante del macrófago (MPIF, del inglés macrophage procoagulant-inducing factor), que permite la expresión de actividad procoagulante del macrófago (MPCA, del inglés macrophage procoagulant activity) en los macrófagos activados; la MPCA también activa la cascada de coagulación, lo que da lugar al depósito de fibrina. El depósito de fibrina es responsable de la induración que se ve en las pruebas cutáneas de HR.
La vía de la HR es importante para eliminar los microorganismos que infectan a los fagocitos. Algunos microorganismos (p. ej., virus) pueden infectar células que carecen de la maquinaría lítica y que no pueden activar la lisis intracelular. Estos agentes patógenos se eliminan mediante CTL. En la infección por un virus, las células expresarán el Ag viral en su superficie asociado con el MHC. Este complejo virus/MHC estimulará la generación de CTL singénicos que lisarán entonces las células que expresan este complejo. Dependiendo de si el producto viral se asocia con la clase I o II del MHC, el CTL pertenecerá a los subgrupos CD8 o CD4, respectivamente. Como se expuso antes, la asociación con cualquiera de las clases de MHC depende de la vía de procesamiento del Ag utilizada; por ejemplo, la mayoría de los CTL generados frente al virus del sarampión y del herpes simple pertenecen al subgrupo CD4. En la infección por el virus de la gripe, los CTL dirigidos frente a Ag nucleoproteico son CD8, mientras que los dirigidos frente al Ag hemaglutinina son CD4.
CÉLULAS B E INMUNIDAD HUMORAL
Las células B constituyen el 5-15% de los linfocitos sanguíneos y son, desde el punto de vista morfológico, indistinguibles de las células T. No obstante, las células B pueden distinguirse por el fenotipo al tener Igs (IgM en las células B inmaduras; IgM e IgD en las células B nativas maduras; IgG, IgA o IgE en las células B que han sufrido cambio de isotipo) y por el CD19, el CD20, el CD21 (CR2), el CD49c, el CD72 y el CD80. Las células B pueden expresar también la clase II del MHC y otros CD que no son específicos de las células B. En los ganglios linfáticos, las células B se encuentran en el área cortical subcapsular externa en los folículos primarios y secundarios y en los cordones celulares; en el bazo, forman la zona marginal y los folículos.
Las células B parece que evolucionan en una serie de pasos programados. Estos pasos comienzan en la médula ósea con la célula primordial comprometida, continúan a través de las fases precoz y tardía de la célula pro-B (con reordenamientos genéticos de las cadenas pesadas D-J) y la fase de célula pre-B (con reordenamiento genético de la cadena pesada V-DJ satisfactorio y aparición de cadenas m citoplásmicas y en la superficie) que finalmente da lugar a la célula B inmadura (con reordenamiento de cadenas ligeras V-J y aparición de IgM en la superficie celular). Se cree que el Ag no interviene en esta secuencia, pero la interacción de las células B inmaduras con el Ag puede provocar inactivación o tolerancia clonal. Las células B inmaduras que no se inactivan pueden seguir evolucionando a células B nativas maduras y dejar la médula para entrar en los órganos linfoides periféricos. Allí, la interacción entre la IgGs y el Ag extraño las convierten en linfoblastos. Al final de la diferenciación, estas células B se convierten en células plasmáticas que secretan Ig de una sola clase.
Las células B de los tejidos periféricos están comprometidas previamente a responder a un número limitado de Ag. La primera interacción Ag-célula B se conoce como respuesta inmunitaria primaria, y las células B comprometidas a responder a este Ag sufren diferenciación y proliferación clonal. Algunas se convierten en células memoria; otras se diferencian a células plasmáticas maduras que sintetizan Ac. Las principales características de la respuesta inmunitaria primaria son un período de latencia antes de la aparición del Ac, la producción sólo de una pequeña cantidad de Ac, inicialmente IgM, y después un cambio de isotipo de Ig (con ayuda de la célula T) a IgG, IgA o IgE. Esto conduce a la creación de muchas células memoria capaces de responder en el futuro al mismo Ag.
La respuesta inmunitaria secundaria (anamnésica o de refuerzo) tiene lugar en encuentros posteriores con el mismo Ag. Las principales características son la proliferación rápida de las células B, la diferenciación rápida a células plasmáticas maduras y la producción rápida de grandes cantidades de Ac, principalmente IgG, que se liberan a la sangre y otros tejidos corporales donde pueden encontrarse y reaccionar de forma eficaz con el Ag.
Frente al mismo Ag pueden generarse IgM, IgG e IgA. De este modo las células B derivadas de una sola célula B nativa madura pueden diferenciarse en una familia de células B genéticamente programadas para sintetizar Ac de una sola especificidad antigénica, con clones representativos comprometidos en la producción de cada clase de Ig (p. ej., IgM, IgG, IgA).
Las células B pueden responder a un Ag de una forma T-dependiente o T-independiente. Los Ag T-independientes (p. ej., polisacáridos neumocócicos, lipopolisacáridos de Escherichia coli y polivinilpirrolidona) tienen un alto peso molecular con determinantes antigénicos que se repiten de una forma lineal y son muy resistentes a la degradación por enzimas del organismo. Los Ag T-independientes provocan sobre todo una respuesta IgM.
La mayor parte de los Ag naturales son T-dependientes y requieren que las células presentadoras de Ag (APC) los procesen. Estas APC presentan el Ag a las células T y a las células B. Las células T liberan citocinas que hacen que la célula B responda al antígeno sintetizando Ac. Durante la estimulación con Ag de las células B, se produce un cambio desde la producción de IgM a la de IgG. Este cambio depende de la célula T cooperadora (TH) y puede necesitar diferentes subgrupos de células TH y citocinas específicas. Por ejemplo, la IL-4 o la IL-13 son necesarias para el cambio de IgM a IgE.
Antígenos y anticuerpos
Estructura y antigenicidad del antígeno. Un Ag es una sustancia que puede desencadenar respuestas inmunitarias específicas. Una vez formados, los Ac pueden combinarse con Ag específicos, igual que las piezas de un rompecabezas. Los Ac reconocen los lugares de combinación del Ag, que son configuraciones específicas (epítopos o determinantes antigénicos) situados en la superficie de las moléculas de alto peso molecular (p.ej., proteínas, polisacáridos y ácidos nucleicos). La presencia de uno de estos epítopos convierte a una molécula en un Ag. Los lugares de combinación del Ag y el Ac se ajustan estrechamente con una intensa fuerza de atracción, porque las áreas emparejadas de la superficie de cada molécula son relativamente grandes. La misma molécula de Ac puede también mostrar reactividad cruzada con Ag relacionados si sus determinantes superficiales son lo suficientemente parecidos a los del Ag original.
Las sustancias son inmunogénicas (antigénicas) si el sistema inmunitario es capaz de reconocer los determinantes antigénicos como extraños (no propios) y si el peso molecular de la sustancia es suficientemente grande. Un hapteno es una sustancia de menor peso molecular que un Ag, que puede reaccionar de forma específica con un Ac, pero que es incapaz de inducir la formación de Ac a no ser que esté unido a otra molécula, habitualmente una proteína (la proteína transportadora); por ejemplo, la penicilina es un hapteno que puede unirse a la albúmina.
Estructura del anticuerpo (v. fig. 146-3). Las moléculas de Ac son Ig que tienen una secuencia de aminoácidos particular y una estructura terciaria para unirse a una estructura complementaria en el Ag. Aunque todas las Ig son probablemente Ac, no siempre es posible saber el Ag al que se dirige cada Ig. La reacción Ag-Ac puede desempeñar un papel específico en la protección del huésped frente a los virus, las bacterias y otros agentes patógenos. La Ig es responsable de la mayor parte de la fracción de g-globulinas de las proteínas del plasma.
Las Ig son muy heterogéneas y en grupo pueden combinarse con un número casi ilimitado de Ag, aunque comparten algunas propiedades comunes. Dentro de cada clase, la Ig monomérica tiene una estructura similar. Cada molécula se compone de cuatro moléculas polipeptídicas: dos cadenas pesadas idénticas y dos cadenas ligeras idénticas. Las cadenas pesadas tienen cada una un peso molecular de unos 50.000-70.000 daltons y las cadenas ligeras de unos 23.000 daltons. Los enlaces disulfuro unen las cadenas y fuerzan a la molécula a la configuración en Y que se conoce habitualmente.
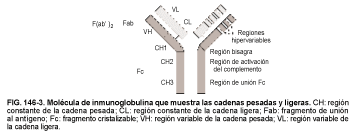
La forma en Y de la molécula de Ig se divide en las regiones variable (V) y constante (C). La región V se localiza en los extremos distales de los brazos de la Y y también se denomina así por la elevada diversidad de aminoácidos que se encuentran aquí, que a su vez determina la capacidad de la Ig de combinarse con el Ag. La región C, proximal al lugar de combinación con el Ag, tiene una secuencia relativamente constante de aminoácidos que es diferente para cada clase de Ig (v. también Inmunidad específica [adaptativa], más arriba).
Las regiones hipervariables dentro de la región V contienen los determinantes idiotípicos, a los cuales pueden unirse los Ac naturales (llamados Ac antiidiotipo). La unión del Ac antiidiotipo con su determinante idiotípico es importante en la regulación de la respuesta de las células B. Por el contrario, los determinantes alotípicos en la región C dan lugar a Ac antialotípicos, que son específicos de clase. De este modo, cada clon de células B produce su propia Ig específica, que tiene una secuencia de aminoácidos específica que se combina con una configuración de Ag particular. No obstante, los miembros de este clon pueden cambiar la clase de molécula de IgE aunque retendrán la cadena ligera y las regiones V.
Las moléculas de Ac se han fragmentado utilizando enzimas proteóliticas para estudiar la relación entre la estructura y la función (v.fig.146-3). La papaína rompe la IgE en dos segmentos monovalentes, el Fab (de unión al Ag), y un solo fragmento Fc (cristalizable). El Fab consta de una cadena ligera y de un fragmento de la cadena pesada y contiene las regiones V de la molécula de Ig (los lugares de combinación). El Fc contiene la mayor parte de la región C; este fragmento es responsable de la activación del complemento y se une a los receptores Fc del complemento. La pepsina produce un fragmento denominado F(ab')2 que consta de dos Fab y de una parte de la cadena pesada con los enlaces disulfuro.
Cada clase de Ig principal en los seres humanos tiene una cadena pesada correspondiente; las cadenas m, g, a, e y d se encuentran en la IgM, IgG, IgA, IgE e IgD, respectivamente. Existen sólo dos tipos de cadenas ligeras, l y k, que se encuentran en las cinco clases humanas de Ig. De este modo, hay 10 tipos diferentes de moléculas de Ig (p. ej., IgG-l, IgG-k). Existen sólo tres clases en forma monomérica (IgG, IgD, IgE). La IgM circula en una forma pentamérica o monomérica. Como pentámero, la IgM contiene cinco moléculas en forma de Y (10 cadenas pesadas y 10 cadenas ligeras). La IgA aparece como un monómero, un dímero y un trímero. La IgG tiene cuatro subclases (IgG1, IgG2, IgG3, IgG4); la IgA tiene dos subclases (IgA1 e IgA2). A las diferentes subclases se les están empezando a asociar funciones biológicas específicas (p. ej., la IgG4 no fija el complemento ni se une a los monocitos, y la IgG3 tiene una semivida significativamente más corta que las otras tres subclases de IgG).
Se han identificado también estructuras adicionales. Las cadenas de unión (J, del inglés joining) unen las cinco subunidades de IgM, así como las subunidades de IgA. La IgA secretora tiene una cadena polipeptídica adicional, el componente secretor (SC), producido por las células epiteliales y que se añade a las moléculas de IgA tras la síntesis de IgA.
De forma tradicional se han utilizado los coeficientes de sedimentación, que se determinan mediante análisis de ultracentrifugación, para denotar cada clase de Ig. La IgM tiene el coeficiente de sedimentación más elevado, de 19S, y la IgG tiene un coeficiente de unos 7S.
Propiedades biológicas de los anticuerpos. La estructura de aminoácidos de la región C de la cadena pesada determina el isotipo de esa clase de Ig. Cada clase sirve para funciones diferentes.
La IgM, el primer Ac formado tras una inmunización primaria (exposición a un Ag nuevo), protege el espacio intravascular de la enfermedad. Las moléculas de IgM pentaméricas activan con facilidad al complemento y sirven como opsonizadoras y aglutinadoras para ayudar al sistema fagocítico a eliminar muchas clases de microorganismos. Las isohemaglutininas y muchos Ac frente a microorganismos gramnegativos son IgM. La IgM monomérica sirve como receptor de Ag en la membrana de la célula B.
La IgG, el tipo más prevalente de Ac sérico, también se encuentra en los espacios extravasculares; se produce cuando los títulos de IgM comienzan a reducirse tras una inmunización primaria. La IgG es la principal Ig que se produce tras la reinmunización (la respuesta inmunitaria de memoria o respuesta inmunitaria secundaria). La IgG protege a los tejidos de las bacterias, los virus y las toxinas, y es la única Ig que atraviesa la placenta. Diferentes subclases de IgG neutralizan toxinas bacterianas, activan el complemento y potencian la fagocitosis mediante la opsonización. La g-globulina comercial es casi por completo IgG, con pequeñas cantidades de otras Ig.
La IgA se encuentra en las secreciones mucosas (saliva, lágrimas, respiratoria, aparato GU y GI y calostro), donde proporciona una defensa precoz antibacteriana y antiviral. La IgA secretora se sintetiza en las regiones subepiteliales de los aparatos GI y respiratorio y se presenta junto con el componente secretor producido a nivel local (SC). En los ganglios linfáticos y en el bazo se encuentran pocas células que produzcan IgA. La IgA sérica no contiene SC. La IgA sérica proporciona protección frente a Brucella, la difteria y la poliomielitis.
La IgD está presente en el suero en concentraciones muy bajas, pero también aparece en la superficie de las células B de desarrollo y puede ser importante en su proliferación y desarrollo.
La IgE (Ac reagínico, sensibilizante cutáneo o anafiláctico), como la IgA, se encuentra sobre todo en las secreciones mucosas respiratorias y GI. En el suero, la IgE está presente en concentraciones muy bajas. La IgE interacciona con los mastocitos; el puenteo de dos moléculas de IgE por el alergeno puede degranular las células, con liberación de mediadores químicos que producen una respuesta alérgica. Las concentraciones de IgE están elevadas en las enfermedades atópicas (p. ej., asma alérgica o extrínseca, fiebre del heno y dermatitis atópica), enfermedades parasitarias, enfermedad de Hodgkin avanzada y mieloma monoclonal de IgE. La IgE puede tener una función beneficiosa frente a los parásitos.
Análisis de las inmunoglobulinas
La IgG, la IgM y la IgA están presentes en el suero en unas concentraciones suficientemente elevadas para que puedan ser cuantificadas mediante diferentes técnicas que miden la presencia de cualquier Ag. La técnica antigua es la inmunodifusión radial (técnica de Mancini), en la que el suero que contiene el Ag se coloca en un pocillo de una placa de agar que contiene el Ac; el tamaño de los anillos de precipitación que se forman en el agar son proporcionales a la concentración del Ag en el suero. Para cuantificar las concentraciones específicas de muchas proteínas séricas, incluidas las Ig, muchos laboratorios utilizan ahora la nefelometría, un método rápido y muy reproducible que utiliza la dispersión molecular de la luz. También se utiliza en ocasiones la inmunoelectroforesis para identificar Ig, sobre todo Ig monoclonal (v. Mieloma múltiple, cap. 140). La IgE está presente en el suero en una cantidad tan pequeña que debe medirse mediante radioinmunoanálisis o análisis inmunoabsorbente ligado a enzima (ELISA). La IgE dirigida frente a un Ag específico se mide utilizando el análisis radioalergoabsorbente (RAST, v. cap. 148). Las subclases de Ig pueden medirse utilizando radioinmunoanálisis o ELISA.
Anticuerpos monoclonales
Los Ac in vivo son casi siempre policlonales (producidos por más de 1 clon), excepto en el caso de la gammapatía monoclonal. De forma similar, los Ac producidos en animales para pruebas diagnósticas han sido policlonales hasta hace muy poco. La técnica del hibridoma permite producir grandes cantidades de Ac monoclonales en animales. Primero, se inmuniza a un ratón con el Ag deseado. Cuando el ratón produce Ac, se le extirpa el bazo para preparar una suspensión de células, algunas de las cuales producen el Ac deseado. Después, estas células productoras de Ac se funden con una línea celular de mieloma que se ha mantenido en cultivo tisular y no produce Ac. Las células individuales fusionadas que producen el Ac monoclonal deseado se aíslan, se hacen crecer en cultivos tisulares para aumentar el número de células y se vuelven a inyectar en el peritoneo del ratón. El líquido ascítico que contiene el Ac monoclonal puede producirse y recogerse con facilidad para obtener concentraciones mayores de Ac. Los laboratorios de experimentación producen preparados comerciales de Ac monoclonales.
Los Ac monoclonales se utilizan ahora ampliamente para 1) medir proteínas y fármacos en el suero; 2) tipificar tejido y sangre; 3) identificar agentes infecciosos; 4) identificar grupos de identificación (CD) para clasificar y seguir las leucemias y los linfomas; 5) identificar Ag tumorales y 6) identificar auto-Ac en diferentes enfermedades. La utilización de los Ac monoclonales ha favorecido la identificación de miríadas de células implicadas en la respuesta inmunitaria.
REGULACIÓN DE LAS RESPUESTAS INMUNITARIAS HUMORALES
La capacidad para montar una respuesta inmunitaria humoral tiene en gran medida una determinación genética. Los genes del MHC regulan el reconocimiento de la célula T del Ag. También son importantes la capacidad de las APC de presentar el Ag y el potencial de la célula B de producir Ac.
El control de la respuesta inmunitaria es crítico. De otra forma, una producción ilimitada de Ac (sobre todo frente a Ag propios) podría provocar una autodestrucción. La respuesta inmunitaria humoral está regulada en primer lugar por la desaparición natural de la sustancia extraña incitadora (p. ej., bacterias) al ser eliminada del organismo. Una regulación adicional viene dada por los Ac y las células T, por la red idiotípica de Ac y por las citocinas. El Ag puede unir el receptor específico del Ag de las células B al receptor de Fcg y, por tanto, suprimir la activación de la célula B nativa. El Ac antiidiotipo reacciona con los determinantes idiotípicos de la región V de la molécula de Ig. Esto se produce porque la región V de cada molécula de Ac es única para el Ac producido por ese clon. A su vez, cada Ac antiidiotípico puede tener idiotipos que serán reconocidos por otros Ac antiidiotípicos, con lo que el proceso de una Ig que reacciona con otra Ig puede continuar. De esta forma, los Ac antiidiotípicos pueden suprimir la producción de los Ac idiotípicos al bloquear los receptores de las células B y T. Este fenómeno explica por qué puede evitarse la enfermedad Rh en el recién nacido mediante la administración pasiva de IgG anti-Rh (anti-D) a la madre.
EL SISTEMA DEL COMPLEMENTO
Un sistema de más de 34 proteínas que interacciona en cascada (similar al sistema de la coagulación) y provoca diferentes procesos biológicos.
Muchas proteínas del complemento son enzimas que existen en el suero como precursores inactivos (cimógenos); muchas otras residen en las superficies celulares. Las proteínas del complemento suponen alrededor del 10% de las proteínas séricas y el tercer componente (C3) está presente en una concentración más alta (alrededor de 1,5 mg/ml). Ver los componentes del sistema del complemento en las tablas 146-2 y 146-3.
Las tres vías de activación del complemento son la clásica, la alternativa y la lectina unida al manana (MBL, del inglés mannan binding lectin) (v. fig. 146-4). Todas se dirigen al paso más importante de la activación, la escisión de C3. La vía final común se denomina vía terminal, o complejo de ataque de membrana (MAC, del inglés membrane attack complex).
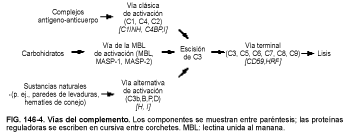

Nomenclatura. Los componentes de la vía clásica se denominan con una C y un número (p.ej., C1, C3). Debido a la secuencia en que fueron identificados, los primeros cuatro componentes se enumeran C1, C4, C2 y C3. Los componentes de la vía alternativa se designan con letras (p.ej., B, P, D). Algunos componentes se denominan factor (p.ej., factor B, factor D). Los componentes o complejos activados tienen una barra sobre ellos para indicar la activación (p.ej., -C1, ---C1r, ----C3b,Bb-). Los fragmentos escindidos se designan con una letra minúscula tras el componentes (p.ej., C3a, C3b son fragmentos del C3). El C3b inactivo se denomina iC3b. Las cadenas polipeptídicas de las proteínas del complemento se designan con una letra griega tras el componente (p.ej., C3a y C3b son las cadenas a y b del C3). Los receptores de membrana celular para el C3 se abrevian CR1, CR2, CR3 y CR4.

La vía clásica
Activación. La vía clásica (v. fig. 146-5) se activa normalmente por Ac fijadores del complemento (Ac que se unen al complemento), que están en complejos Ag-Ac o en los cuales se agrega el Ac (IgG o IgM). De este modo, la vía clásica sirve a la inmunidad específica porque sólo el Ac de clases específicas, formado en respuesta a la estimulación por el Ag, es capaz de activar esta vía. La macromolécula C1 es un complejo de una molécula de C1q, dos de C1r y dos de C1s que dependen de Ca++. La macromolécula C1 permanece intacta sólo cuando hay Ca++ presente; de otra forma las unidades individuales se disociarían entre sí. La activación se produce cuando dos de los seis monómeros del C1q se unen a las regiones Fc de las moléculas de IgG o a una molécula de IgM pentamérica. Debe haber dos moléculas de IgG adecuadamente preparadas para que se produzca la activación, mientras que una sola IgM pentamérica contiene esa proximidad en su estructura. Por tanto, la IgM es mucho más eficaz para activar el complemento que la IgG. La actividad de la IgG es en orden IgG3 > IgG1 > IgG2. La IgG4 no fija el complemento.
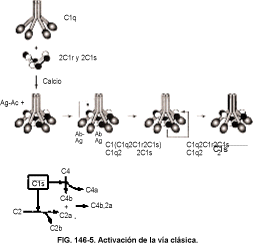
Una vez que la Ig se une al C1q, la molécula de C1q cambia su estructura terciaria y provoca la activación autocatalítica de C1r a --C1r. El C1r escinde entonces un enlace de C1s para producir --C1s. No se libera ningún fragmento cuando se escinden C1r o C1s.
El --C1s también se denomina esterasa de C1. El --C1s puede escindir C4a y C4b. El C4b, el principal fragmento de escisión, se une a la membrana si está presente. El --C1s puede entonces liberar C2 libre para producir C2a y C2b, que es un proceso ineficaz, o escindir C2 en un complejo C4b,C2 para producir C4b, C2a y C2b libre, que es un proceso muy eficaz. El C2a es el principal fragmento de escisión de C2. Si se ha escindido C2 libre, entonces C2a debe unirse a C4b para formar C4b,2a, o el C2a se degrada e inactiva. El C4b,2a es la convertasa del C3 de la vía clásica, que puede escindir el C3 en C3a y C3b. El C2a contiene el sitio enzimático donde se produce la escisión del C3. El C4b,2a necesita la presencia de magnesio y se degrada con el tiempo a temperatura fisiológica.
La vía clásica también puede activarse por mecanismos independientes del Ac. La heparina (un anticoagulante polianiónico) y la protamina (un policatión que se utiliza para bloquear la heparina), cuando están presentes en concentraciones equimolares, pueden activar la vía clásica. Se cree que otros polianiones (p. ej., ADN y ARN) pueden activar la vía clásca. La proteína C reactiva es capaz de activar la vía clásica sin la presencia de Ac. Se han descrito vías que eluden el C1 y que no utilizan los componentes de la vía clásica pero escinden el C3. Una de ellas se ha caracterizado como la vía MBL.
Regulación. La vía clásica la regula el inhibidor de la esterasa de C1 (C1INH), que se une de forma estoiquiométrica (1:1) al C1r y C1s y al --C1r y --C1s para inactivar estas proteínas de forma permanente. El C1INH también se une de forma estoiquiométrica a la plasmina, la calicreína, el factor de Hageman activado y el factor XIa de la coagulación. Su ausencia provoca el angioedema hereditario (v. cap. 148). El factor J es una glucoproteína catiónica que también inhibe la actividad del C1. La proteína de unión al C4 (C4BP, del inglés C4-binding protein) rompe el complejo C4b,2a, lo que permite al factor I inactivar al C4b.
La vía alternativa
Activación. La vía alternativa (v. fig. 146-6) la activan sustancias naturales (p. ej., paredes de levaduras, factor de veneno de cobra, factor nefrítico, pared de células bacterianas [endotoxina], hematíes de conejo [in vitro]) y la IgA agregada como respuesta inmunitaria inespecífica (innata), es decir, una que no requiere sensibilización previa. En la vía alternativa no intervienen el C1, el C4 ni el C2, pero sí escinde el C3. Esta vía depende de la escisión constante de pequeñas cantidades de C3 en C3a y C3b. Se sabe poco de esta escisión natural del C3 y se cree que ocurre por medio de una acción inespecífica de las enzimas sobre el C3 o por una actividad de nivel bajo de las otras dos vías. El C3b sirve entonces de sustrato del factor B para producir el complejo C3b,B. La properdina (P) estabiliza este complejo C3b,Bb retrasando su degradación. El C3b,Bb y el C3b,Bb,P son las convertasas del C3 de la vía alternativa, las enzimas que escinden el C3 en C3a y C3b. El Bb contiene los lugares enzimáticos para escindir el C3. El C3b,Bb necesita la presencia de magnesio y se degrada con el tiempo.
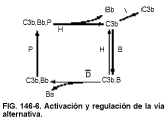
La vía alternativa se ve como una vía de amplificación porque un complejo C3b,Bb puede escindir muchas moléculas de C3. Sin embargo, también se produce la amplificación cuando se produce --C1s y cuando se forma C4b,2a. Cada una de estas enzimas puede escindir cientos de moléculas, provocando una activación rápida del complemento.
Regulación. El complejo C3b,Bb de la vía alternativa está regulado por varios factores. La properdina retrasa la degradación del complejo C3b,Bb, aumentando su semivida de unos 4 min a 40 min. Las sustancias aceleradoras de la degradación (p. ej., el factor H o factor acelerador de la degradación [DAF, del inglés decay accelerating factor]) compiten con B para unirse al C3b (p. ej., para producir C3b,H), reduciendo la semivida del complejo C3b,Bb y provocando la disociación del complejo en C3b y Bb. El factor I actúa sobre el C3b,H para degradar el C3b (lo que provoca la producción de iC3b, C3c, C3d, C3f y C3dg).
Las circunstancias bajo las cuales se forma el complejo C3b,Bb determinarán si se activa o no la vía alternativa. Las superficies a las que el complejo C3b,Bb puede unirse son superficies activadoras (p. ej., paredes de levaduras, hematíes de conejo) o no activadoras (p. ej., hematíes de carnero). Las superficies activadoras evitan que el factor H se una al C3b, mientras que las superficies no activadoras permiten que el factor H se una al C3b y disocie el C3b,Bb. Por tanto, el complejo C3b,Bb permanece activo mucho más tiempo en una superficie activadora que en una no activadora.
Los mecanismos descritos antes explican cómo la vía alternativa se activa in vivo. El factor de veneno de cobra (CoVF) es similar al C3b de cobra; el complejo CoVF,Bb es muy estable y no es susceptible a la acción degradadora del factor H. Por tanto, el CoVF,Bb puede escindir de forma rápida y casi total el C3. El factor nefrítico C3 (C3NeF) se encuentra en el suero de aproximadamente el 10% de los pacientes con glomerulonefritis membranoproliferativa y es una Ig dirigida frente al complejo C3Bb. El C3NeF actúa como la properdina, excepto en que el complejo C3b,Bb,B3NeF es relativamente resistente a la actividad degradadora del factor H. Las paredes de levaduras (cimosán) y ciertas membranas (p.ej., hematíes de conejo) son superficies activadoras sobre las cuales el complejo C3b,Bb está protegido de la actividad degradadora del factorH.
Vía de la lectina unida al manana
La vía de la lectina unida al manana (MBL) depende del reconocimiento innato de sustancias extrañas (es decir, carbohidratos) para la activación. Esta vía tiene similitudes estructurales y funcionales con la vía clásica. La MBL es similar al C1q y el MASP-1 y el MASP-2 parecen ser similares al C1r y C1s de la vía clásica, respectivamente. De este modo, el MASP-2 puede escindir el C4 y conducir a la formación de una convertasa del C3 derivada de la vía de la MBL.
Escisión del C3 y sus consecuencias
Las convertasas del C3 escinden el C3 en C3a y C3b, que genera un lugar de unión metaestable en el C3b para las membranas. Si se dispone de una superficie o una membrana inmediatamente después de que la convertasa del C3 actúe sobre el C3, el C3b puede unirse de forma covalente. Si no se dispone de una membrana o superficie, entonces el C3b se convierte en C3b de fase fluida y es incapaz de unirse de forma covalente a las superficies celulares. El C3 también puede convertirse en una molécula similar al C3b si se trata con metilamina. Una vez que el C3b se ha unido a la membrana a través del lugar de unión metastable lábil, puede participar en las actividades biológicas al unirse a diferentes receptores del C3, servir como lugar de unión eficaz para que B escinda más C3 a través de la vía alternativa, participar en la formación de la convertasa del C5 o ser activado por el factor I y un cofactor para formar iC3b.
De este modo, el C3b puede unirse de forma covalente a las membranas por su lugar de unión metaestable tioléster y una vez unido puede interaccionar con diferentes receptores dependiendo de la disponibilidad de receptores del C3 en las células y del estado de degradación del C3. La unión a las membranas a través del lugar de unión metaestable covalente no debe confundirse con la unión no covalente a los receptores.
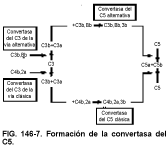
Complejo de ataque de la membrana: la vía terminal
La convertasa del C3 (p. ej., C3b,b) puede convertirse en convertasa del C5 (p. ej., C3b,Bb,3b) por la adición de un C3b al complejo (v. fig. 146-7). La convertasa del C5 escinde C5 en C5a y C5b, comenzando la formación del complejo de ataque de la membrana (MAC, del inglés membrane attack complex). El C6 puede entonces unirse al C5b para producir C5b,6. Después, el C7 puede unirse para formar C5b,6,7, que puede unirse a las membranas y a las bicapas lipídicas. Cuando esto se produce en una célula que no tiene ningún producto del complemento sobre ella, se habla del fenómeno del espectador inocente (y puede causar la hemólisis de la célula inocente). El C8 puede entonces unirse al complejo C5b,6,7 para formar C5b,6,7,8, que puede provocar una lisis lenta e ineficaz de la célula. Finalmente, el C9 se une al complejo para producir C5b,6,7,8,9, que inicia una lisis sustancial de la célula. Como se añaden moléculas de C9 adicionales al complejo C5b-9, la lisis aumenta. El MAC está regulado por la proteína S, también denominada vitronectina (que controla la actividad de C5b-7), por el factor de restricción homólogo (HRF), por el SP40,40 y por el CD59 (que regula la actividad de C8,9).
Actividades biológicas asociadas con la activación del complemento
La lisis celular es sólo una de las muchas actividades biológicas asociadas con la activación del complemento y puede no ser la más importante. La lisis se ve a nivel clínico en los pacientes con hemoglobinuria paroxística nocturna, una enfermedad rara debida a un déficit de las proteínas de membrana DAF (factor acelerador de la degradación), HRF (factor de restricción homólogo) y CD59.
Los receptores del complemento están presentes en diferentes células. El CR1, la proteína cofactor de membrana (MCP, CD46) y el DAF (CD55) regulan la degradación del C3b. El HRF y el CD59 evitan la formación del complejo de ataque de la membrana en células homólogas. El CR1 (CD35) también interviene en el aclaramiento de complejos inmunitarios. El CR2 (CD21) regula las funciones de la célula B (producción de Ac) y es el receptor del virus de Epstein-Barr. El CR3 (CD11b/CD18) interviene en la fagocitosis al mediar la adhesión de partículas cubiertas de iC3b para la fagocitosis. El CR4 aparece en las plaquetas y ha sido menos estudiado que los otros receptores del C3. El gp150,95 interviene en la migración del monocito. Los receptores del C3a y C4a se unen al C3a y C4a, respectivamente. El receptor de C5a se une al C5a y al C5adesarg (C5a sin la arginina terminal) que está presente en una amplia variedad de células. El receptor del C1q se une a la porción colágena del C1q, lo que permite la unión de complejos inmunitarios a los fagocitos.
El C3a y el C5a tienen actividad anafilotoxina y el C4a tiene una actividad de anafilotoxina débil. Las anafilatoxinas aumentan la permeabilidad vascular, contraen el músculo liso y degranulan el mastocito. Las anafilatoxinas están reguladas por el inactivador de la anafilatoxina (N carboxipeptidasa), que en segundos eliminan la arginina carboxiterminal.
La quimiotaxis es la atracción de las células a un área de inflamación. El C5a tiene actividad de anafilotoxina y quimiotáctica, pero el C3a y el C4a no son quimiotácticos. También se ha descrito quimiotaxis con el iC5b-7.
El C5a y el C5adesarg regulan las actividades del neutrófilo y del monocito. El C5a puede aumentar la adherencia de las células, provocar la degranulación y liberación de enzimas intracelulares a partir de los granulocitos, producir O2 tóxico e iniciar otros acontecimientos metabólicos celulares.
El aclaramiento de complejos inmunitarios es una función importante del complemento. La vía clásica puede evitar que se formen grandes complejos inmunitarios y la vía alternativa puede aumentar la solubilidad de los complejos inmunitarios.
Las proteínas del complemento también pueden tener otras actividades biológicas. Los fragmentos de C3 (C3d o C3dg) pueden ayudar a regular la producción de Ac a través del CR2 de las células. El angioedema hereditario, que está causado por un déficit de inhibidor de C1, puede estar mediado por una sustancia similar a la cinina poco descrita. Un fragmento mal definido del C3 (C3e, factor movilizador de leucocitos) puede movilizar leucocitos de la médula ósea. El fragmento Bb del factor B aumenta la diseminación y adherencia de los macrófagos. La activación del complemento puede también neutralizar a los virus y provocar leucocitosis.
Análisis de la actividad funcional del complemento
El análisis del complemento hemolítico total (CH50) mide la capacidad de la vía clásica y del MAC de lisar hematíes de carnero a los cuales se ha unido un Ac. El CH50 de la vía alternativa (CH50 de conejo o APCH50) mide la capacidad de la vía alternativa y del MAC de lisar hematíes de conejo. Los análisis hemolíticos pueden utilizarse para medir la actividad funcional de componentes específicos de cualquier vía. Las proteínas del complemento también pueden medirse utilizando técnicas antigénicas (p. ej., nefelometría, difusión en gel de agar, inmunodifusión radial).
El complemento también puede utilizarse como reactivo para ayudar en el diagnóstico. En la prueba de fijación del complemento se calienta el suero del paciente para destruir las enzimas del complemento. Después, se añade Ag (p.ej., una partícula viral) y complemento adicional al suero del paciente y la mezcla se incuba. Finalmente, se añaden hematíes de carnero y se continúa la incubación. Si el sistema del complemento se ha activado por la presencia del Ac en el suero del paciente, la actividad hemolítica del complemento estará gastada y el hematíe no se lisará. Si no hay Ac en el suero del paciente, los hematíes se lisarán.
RESOLUCIÓN DE UNA RESPUESTA INMUNITARIA
Una respuesta inmunitaria puede asociarse con una proliferación y diferenciación masiva de linfocitos (p. ej., aumento del tamaño de las amígdalas y dolor faríngeo). ¿Qué le sucede a los linfocitos cuando la infección se controla? Como se dijo antes, la respuesta inmunitaria se asocia con la secreción de varias citocinas. Cuando la infección está controlada y los Ag se han eliminado, la secreción de citocinas se detiene. Cuando la secreción de citocinas cesa, los linfocitos sufren apoptosis. Las células mueren de dos formas.
1. La necrosis se refiere a los cambios morfológicos que se producen cuando una célula muere por una lesión intensa y brusca (p. ej., lisis osmótica, isquemia, hipertermia, traumatismo químico). La mayor parte de la lesión se produce en la membrana plasmática, lo que provoca la pérdida de la capacidad de la célula de regular la presión osmótica y da lugar a su vez a la ruptura de la célula y al vertido de su contenido en el tejido circundante. Esto origina una respuesta inflamatoria.
2. La apoptosis (también denominada muerte celular programada) es muy común en los invertebrados. Por ejemplo, cuando una mariposa sale de su capullo ya no necesita más los músculos que utilizó para ese proceso; estos músculos sufren una muerte celular programada. En los mamíferos, la apoptosis se refiere al proceso por el cual una célula «se compromete a suicidarse» y se caracteriza por una serie de cambios morfológicos. La apoptosis comienza por la condensación de la cromatina (secundaria a la activación de una endonucleasa endógena que degrada el ADN) y la rotura del núcleo colapsado en pequeños fragmentos. A la vez se produce la zeiosis (formación de burbujas en la membrana plasmática) que puede servir como señal para la fagocitosis por los macrófagos vecinos. Al contrario de lo que ocurre en la necrosis, esta fagocitosis inmediata evita que se derrame el contenido celular y el desarrollo de inflamación.
La apoptosis es un proceso activo e implica la inducción de varias moléculas y vías. Dos vías implicadas en la apoptosis ilustran las posibles enfermedades que pueden producirse por una apoptosis normal. Una enzima llamada Bcl-2 puede inhibir la apoptosis. Por tanto, si se induce a un linfocito a expresar Bcl-2 no morirá y persistirá; esto es lo que sucede en algunos linfomas (Bcl significa linfoma de célula B, que es donde se descubrió por primera vez la Bcl-2). En la apoptosis también participa una interacción molécula-ligando en la superficie celular. Muchas células expresan Fas (CD95) en su superficie. El entrecruzamiento del Fas activa la vía de la apoptosis. Éste es un mecanismo importante por el cual el CTL lisa sus célula dianas al adquirir el ligando del Fas; el ligando del Fas se une al Fas en la célula diana y provoca la activación de la apoptosis en la célula diana. La ausencia del Fas o del ligando del Fas puede, en teoría, provocar la persistencia de los linfocitos y una linfadenopatía masiva. Esto ocurre en modelos animales donde un déficit de Fas (ratones lpr) o del ligando del Fas (ratones gld) provoca una linfadenopatía masiva y autoinmunidad. Las anomalías en el Fas también se han comunicado en seres humanos y son la causa subyacente del síndrome de Canale-Smith.
Ciertos órganos (p. ej., la retina, los testículos) son «lugares privilegiados» en el sentido de que el sistema inmunitario los ignora o tolera. Ahora parece que estos órganos expresan una elevada densidad de ligando del Fas en su superficie celular. Cualquier linfocito que trate de atacar a estos órganos sufrirá un entrecruzamiento del Fas y el propio linfocitos sufrirá apoptosis. Esta estrategia de evasión inmunitaria también la emplean muchos tumores; algunos tumores expresan el ligando del Fas en su superficie y por ello inducen la apoptosis de cualquier linfocito que intente atacarles.