234 / ENDOCRINOLOGÍA DE LA REPRODUCCIÓN
La función reproductora normal requiere una compleja interacción de los órganos endocrinos y los órganos diana. La función normal es esencial para el desarrollo sexual puberal y de los procesos cíclicos de ovulación y menstruación.
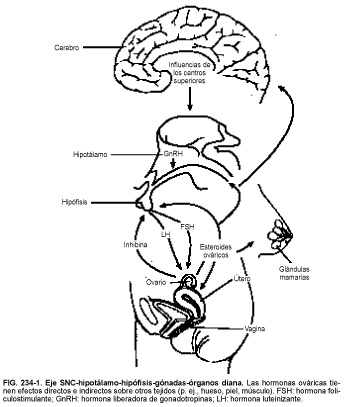
El hipotálamo segrega un pequeño péptido, hormona liberadora de gonadotropinas (GnRH), también conocida como hormona liberadora de hormona luteinizante, que regula la liberación de las gonadotropinas hormona luteinizante (LH) y hormona foliculoestimulante (FSH) de la hipófisis anterior (v.fig. 234-1 y caps. 6 y 7). La LH y la FSH promueven la maduración de las células germinales y estimulan la secreción ovárica de estrógenos y progesterona.
Estrógenos y progesterona son compuestos policíclicos (con los átomos de carbono formando cuatro anillos) derivados del colesterol. Estas hormonas circulan por la sangre unidas casi por completo a varias proteínas plasmáticas. Sólo los estrógenos y la progesterona no unidos a proteínas parecen ser biológicamente activos. Estimulan los órganos diana del aparato reproductor (es decir, mama, útero y vagina) y ejercen efectos de retroacción negativa y positiva en el eje SNC-hipotálamo-hipófisis, inhibiendo y estimulando la secreción de gonadotropinas.
Virtualmente todas las hormonas se liberan en brotes cortos o pulsos a intervalos de 1 a 3 h, con lo que los patrones descritos son meras representaciones idealizadas. Estos factores se deben tener en cuenta al interpretar valores hormonales puntuales.
Secreción hormonal: de la infancia a la pubertad
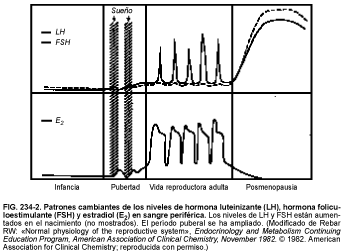
Tanto la LH como la FSH están elevadas en el momento del nacimiento, pero descienden hasta niveles bajos al cabo de algunos meses, manteniéndose así a lo largo de los años prepuberales, siendo los niveles de FSH ligeramente más elevados que los de LH (v. fig. 234-2).
Los andrógenos suprarrenales, deshidroepiandrosterona (DHEA) y su derivado sulfato (DHEAS), comienzan a aumentar varios años antes de la pubertad. Este aumento puede ser importante en el inicio del crecimiento del vello púbico y axilar (es decir, la adrenarquia) y otros fenómenos de la pubertad. Dado que la ACTH y el cortisol no aumentan con estos andrógenos, se ha sugerido que otro péptido hipofisario, aún no identificado, inicia la secreción de andrógenos suprarrenales.
No se conocen con exactitud los mecanismos responsables del inicio de la pubertad. Influencias centrales pueden inhibir la liberación pulsátil de GnRH durante la infancia, iniciando posteriormente su liberación para inducir la pubertad en los primeros años de la adolescencia.
Al principio de la pubertad hay un descenso de la sensibilidad del hipotálamo a los esteroides gonadales, aumentándose la secreción de LH y FSH y estimulándose la producción de esteroides gonadales (principalmente estrógenos) y el desarrollo de las características sexuales secundarias. Al principio, el incremento en la secreción de LH y FSH se produce sólo durante el sueño, aumentando después todo el día. Los patrones de incremento de los niveles basales de LH y FSH son diferentes para los niños y las niñas pero los incrementos de LH son superiores en ambos a los de FSH.
Pubertad
Secuencia de acontecimientos por la que un niño alcanza el crecimiento y la madurez sexual.
Los cambios físicos de la pubertad se producen secuencialmente durante la adolescencia. El desarrollo de las mamas en las niñas suele ser el primer cambio puberal, seguido inmediatamente por la aparición de vello púbico y axilar (v. fig. 234-3). El intervalo desde el inicio del desarrollo de las mamas hasta la menarquia (primera menstruación en la mujer) suele ser de unos 2 años. El brote de crecimiento de las adolescentes que acompaña a la pubertad comienza típicamente antes del inicio del desarrollo mamario, pero raras veces se identifica. Las niñas alcanzan la velocidad máxima de crecimiento precozmente en la pubertad, antes de la menarquia, y después de ésta tienen un potencial de crecimiento limitado. El hábito cambia, aumentando el porcentaje de grasa corporal.
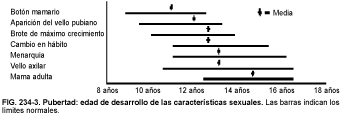
La edad en la cual comienza la pubertad es variable y parece estar influida por el estado general de salud y nutrición, la condición socioeconómica y factores genéticos. En las naciones industrializadas, la edad del inicio de la pubertad ha descendido de manera importante; por ejemplo, en Europa Occidental se ha producido un descenso en la edad de la menarquia de 4 meses por cada década entre 1850 y 1950, aunque no ha descendido en las cuatro últimas décadas. La obesidad moderada se asocia a una menarquia precoz, mientras que la menarquia retardada es común en niñas con un peso muy inferior al normal o malnutridas. Estas observaciones han llevado a postular la teoría de que se debe alcanzar un peso corporal crítico para que ocurra la menarquia. También se produce un desarrollo puberal precoz en las niñas que viven en áreas urbanas, las niñas ciegas y aquellas cuyas madres tuvieron una madurez sexual temprana.
Desarrollo folicular ovárico
En la 6.ª sem de gestación, las células germinales primordiales (oogonias) han migrado mediante movimientos ameboides desde su localización de origen en el saco vitelino a las crestas genitales (presuntos ovarios). Su número aumenta notablemente por mitosis durante el 4.º mes de la gestación, tras el cual la mayoría sufre atresia. Durante el 3er mes, algunas células comienzan a dividirse por meiosis en lugar de mitosis, y para el 7.º mes todas las células viables queden detenidas en la fase de diploteno de la profase meiótica; estas células se denominan oocitos. Entre el 7.º y el 9.º mes de gestación, el ovario fetal se organiza y cada oocito pasa a formar parte de un folículo primordial, constituido por una membrana basal, una capa única de células epiteliales escamosas de la granulosa y un oocito. Los folículos primordiales constituyen la reserva de folículos que están en reposo, y se constituirán en la reserva de crecimiento (de la que se desarrollarán todos los folículos maduros) o sufrirán atresia. Los mecanismos que inician el crecimiento folicular y del oocito no están dilucidados, pero se sabe que no se requiere la participación de las gonadotropinas.
La hembra humana nace con un número limitado de óvulos, 99,9% de los cuales se eliminarán por atresia. Como cada oocito permanece detenido en la profase meiótica hasta que es ovulado, es una de las células del cuerpo de vida más larga (desde embrión hasta alrededor de los 50 años). La dilatada duración de la vida de los oocitos puede explicar el aumento de la incidencia de gestaciones genéticamente anormales a medida que aumenta la edad de la madre.
Durante los años reproductores de una mujer, varios folículos incluidos en la reserva de crecimiento son reclutados en cada ciclo, siendo generalmente uno el seleccionado para la ovulación (v. fig. 234-4). Este folículo da lugar al folículo de De Graaf (preovulatorio), que es capaz de responder al aumento de LH de la mitad del ciclo. Se desconoce el mecanismo de selección implicado.
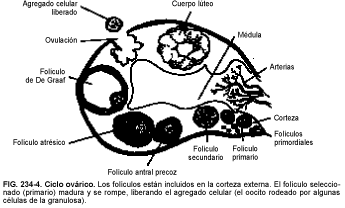
El folículo de De Graaf contiene un antro (cavidad rellena de líquido), creada por la proliferación de células de la granulosa, que segregan líquido y mucopolisacáridos. El aumento del tamaño folicular se debe principalmente a la acumulación de líquido folicular, controlado por la FSH, induciéndose igualmente la proliferación de receptores específicos para la LH en las células de la granulosa. Los receptores de la LH son responsables de la estimulación de la secreción de progesterona que ocurre antes de la ovulación y de la producción continua de progesterona en la fase lútea. Las células de la granulosa del folículo también desarrollan receptores de membrana específicos para prolactina, que descienden en número a medida que madura el folículo; se desconoce su papel fisiológico.
Ciclo menstrual
La menstruación es la pérdida vaginal debida a la descamación del endometrio, que se produce con un intervalo aproximado de un mes, durante la vida reproductora de una mujer; esta pérdida sanguinolenta se denomina período o flujo menstrual.
El primer día de la menstruación es el día 1 del ciclo menstrual. La duración media de la menstruación es de 5 (±2) d. Aunque la media de la duración del ciclo menstrual es de 28 d, sólo del 10 al 15% de los ciclos normales tiene exactamente 28 d de duración; el rango normal para un ciclo ovulatorio es de entre 25 y 36 d. La variación máxima de los intervalos intermenstruales se produce generalmente en los años que siguen a la menarquia y en los que preceden a la menopausia, cuando son más comunes los ciclos anovulatorios. La pérdida sanguínea es de unos 130 ml por ciclo (rango 13-300 ml), siendo normalmente más copiosa en el 2.º día. Una compresa o un tampón pueden absorber de 20 a 30 ml. Habitualmente la sangre menstrual no se coagula (a no ser que el sangrado sea muy copioso), probablemente debido a la presencia de fibrinolisina y de otros factores inhibidores de la coagulación.
Basándose en los acontecimientos endocrinos, el ciclo menstrual se puede dividir en 3 fases (v.fig. 234-5). La fase folicular (preovulatoria) comienza el primer día de sangrado y se extiende hasta el día anterior al aumento preovulatorio de la LH; es la fase más variable en cuanto a duración. En la primera mitad de esta fase, aumenta ligeramente la secreción de FSH, estimulando el crecimiento de un grupo de 3 a 30 folículos que han sido reclutados debido a su crecimiento acelerado durante los últimos días del ciclo precedente. Conforme descienden los niveles de FSH, se selecciona uno de estos folículos para ovular; éste madura y los demás sufren atresia. Los niveles circulantes de LH aumentan lentamente, a partir de 1-2 d tras la elevación de la FSH. La secreción de estrógenos y progesterona por los ovarios es relativamente constante y se mantiene baja en este período.
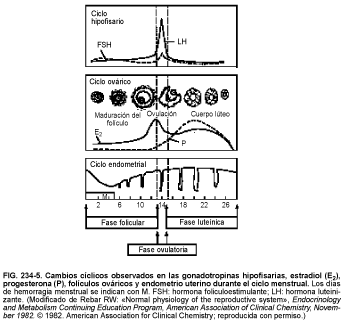
Unos 7-8 d antes de la oleada preovulatoria, la secreción estrogénica ovárica del folículo seleccionado, en particular estradiol, aumenta, lentamente al principio y luego con rapidez, hasta alcanzar su pico, generalmente en el día previo a la oleada de LH. Este aumento de estrógenos se acompaña de un aumento lento pero constante de los valores de LH y de una disminución de los de FSH. La divergencia entre los valores de LH y de FSH puede deberse a la acción inhibidora sobre la FSH de los estrógenos (comparada con la liberación de LH) junto con la inhibición específica por la inhibina. Antes de la oleada de LH, los valores de progesterona empiezan a aumentar de forma significativa.
En la fase ovulatoria, una serie de complejos acontecimientos endocrinos culminan en la oleada de LH: la liberación masiva preovulatoria de LH por la hipófisis. La liberación ovulatoria de LH se produce, en parte, como consecuencia de una retroacción estrogénica positiva. Se produce simultáneamente un incremento menor de la secreción de FSH, cuyo significado se desconoce. Al aumentar los valores de LH, bajan los de estradiol, pero las concentraciones de progesterona siguen aumentando. La oleada de LH dura típicamente 36-48 h y consta de múltiples y amplias oleadas de LH liberadas de forma pulsátil. La oleada de LH, que da lugar a la completa madurez del folículo, es necesaria para la ovulación (liberación del oocito desde el folículo de De Graaf maduro) que ocurre de 16-32 h después del inicio de la oleada. Se desconoce el mecanismo que causa la ovulación.
Durante el aumento de la LH, el folículo preovulatorio se hincha y protruye en el epitelio ovárico. Aparece un estigma o mancha avascular en la superficie del folículo. Se forma una pequeña vesícula en el estigma, la cual se rompe, y el agregado (oocito y algunas células de la granulosa que lo rodean) es expulsado. La producción de prostaglandinas por el mismo folículo, tal vez regulada por la LH y/o FSH, también parece esencial en el proceso ovulatorio. Las enzimas proteolíticas de las células de la granulosa y de las células epiteliales que se encuentran sobre el folículo preovulatorio, factores de crecimiento locales y citocinas parecen desempeñar un papel importante en la rotura del folículo. El oocito permanece frenado en la profase meiótica hasta el momento posterior al pulso de LH. Dentro de las 36 h siguientes a la oleada de LH, el oocito completa su 1.ª división meiótica, en la que cada célula recibe 23 cromosomas de los 46 originales y es expulsado el primer corpúsculo polar. En la segunda división meiótica, en la cual cada cromosoma se divide longitudinalmente en pares idénticos, no se completa ni se elimina el 2.º corpúsculo polar a menos que el huevo sea penetrado por un espermatozoide.
En la fase lútea (postovulatoria), las células de la teca y la granulosa, que constituyen el folículo, se reorganizan para formar el cuerpo lúteo (cuerpo amarillo), que da nombre a esta fase. La duración de esta fase es la más constante, un promedio de 14 d en ausencia de embarazo, finalizando con el 1.er d de la menstruación. La duración de esta fase proviene de la duración de la vida funcional del cuerpo lúteo, que segrega progesterona y estradiol durante unos 14 d, degenerando posteriormente si no se produce la fertilización. El cuerpo lúteo mantiene la implantación del óvulo fecundado, segregando progesterona en cantidades crecientes, alcanzando un pico de 25 mg/d en los 6 a 8 d posteriores a la oleada de LH. Debido a que la progesterona es termogénica, la temperatura basal aumenta en por lo menos 0,5°C en la fase lútea y se mantiene elevada hasta la menstruación. Las prostaglandinas y el IGF-II pueden desempeñar un papel en la regulación del tiempo de vida del cuerpo lúteo; sin embargo, poco se sabe sobre ello en la actualidad.
Si se produce la fertilización, la gonadotropina coriónica humana (HCG) del óvulo fecundado mantiene al cuerpo lúteo hasta que la unidad fetoplacentaria puede mantenerse a sí misma endocrinológicamente. La hCG es estructural y funcionalmente similar a la LH; sin embargo, de forma típica, las pruebas de embarazo utilizan anticuerpos específicos para la subunidad b de la hCG y tiene pocas reacciones cruzadas, o ninguna, con la LH.
Durante casi toda la fase lútea, los niveles de LH y FSH circulantes disminuyen y se mantienen bajos durante la mayor parte de la fase lútea, pero comienzan a aumentar nuevamente con la menstruación (ciclo siguiente).
Cambios cíclicos en otros órganos del aparato reproductor
Endometrio. Los cambios cíclicos que se producen en el endometrio culminan en la menstruación. El endometrio, compuesto de glándulas y estroma, tiene tres capas: la capa basal, la capa esponjosa intermedia y la capa superficial de células epiteliales compactas, que cubre la cavidad uterina. La capa basal no se pierde durante las menstruaciones y regenera las otras dos capas que si se pierden. Dado que los cambios histológicos que se producen durante el ciclo menstrual son característicos, se pueden utilizar las biopsias endometriales para determinar con exactitud la fase del ciclo y valorar las respuestas tisulares a los esteroides gonadales.
Al inicio de la fase folicular, el endometrio es delgado (aproximadamente 2 mm), con glándulas endometriales estrechas y rectas con un epitelio columnar bajo. El estroma es denso. A medida que aumentan los niveles de estradiol, durante el final de la fase folicular, el endometrio crece rápida y progresivamente con extensas mitosis (es decir, regeneración a partir de la capa basal) hasta los 11 mm; la mucosa se engruesa, y las glándulas tubulares se alargan, transformándose en tortuosas. El endometrio puede verse mediante ecografía transvaginal; éste posee en esta fase un patrón trilaminar característico, que se transforma en homogéneo tras la ovulación.
Durante la fase lútea, y bajo la influencia de la progesterona, las glándulas tubulares se dilatan, se llenan con glucógeno y se transforman en secretoras con el aumento de la vascularización del estroma. A medida que los niveles de estradiol y progesterona disminuyen al final de la fase luteínica, el estroma se vuelve edematoso, hay necrosis de los vasos sanguíneos y del endometrio y se produce la hemorragia endometrial.
Cuello uterino. Durante la fase folicular se produce un aumento progresivo de la vascularización cervical, de la congestión, del edema y de la secreción de moco cervical. El orificio cervical externo se abre hasta alcanzar un diámetro de 3 mm en la ovulación, reduciéndose posteriormente a 1 mm. El incremento progresivo de los estrógenos provoca un aumento de 10-30 veces en la cantidad de moco cervical. Las características del moco cervical son útiles clínicamente para evaluar la etapa del ciclo y el estado hormonal de la paciente. La elasticidad (filancia) aumenta y la arborización (imágenes en hojas de helecho que se obtienen cuando se examina microscópicamente una muestra de moco cervical secado sobre un portaobjetos de vidrio) se hace evidente justo antes de la ovulación. Esta imagen es el resultado de un aumento de NaCl en el moco cervical por la influencia estrogénica. Durante la fase luteínica, bajo la influencia de progesterona, el moco cervical se espesa, se hace menos acuoso y pierde su elasticidad y capacidad para formar arborizaciones en helecho.
Vagina. La proliferación y la maduración del epitelio vaginal también se ven influidas por los estrógenos y la progesterona. Cuando al inicio de la fase folicular la secreción de estrógenos ováricos es baja, el epitelio vaginal es delgado y pálido. A medida que aumentan los niveles de estrógenos en la fase folicular, las células escamosas maduran y se cornifican, engrosándose el epitelio. Durante la fase luteínica, el número de células intermedias precornificadas aumenta, y se incrementa el número de leucocitos y detritos a medida que se van desprendiendo las células escamosas maduras. Los cambios del epitelio vaginal pueden cuantificarse histológicamente y utilizarse como índice cualitativo de la estimulación estrogénica.
Regulación neuroendocrina del ciclo menstrual
La secreción pulsátil de LH y FSH está determinada por la secreción pulsátil de GnRH. La frecuencia y amplitud de los pulsos de LH y FSH está modulada por los esteroides ováricos y varía a lo largo del ciclo menstrual. No se ha identificado una hormona liberadora diferente para la FSH. Hay evidencias que sugieren que las mismas células a veces contienen LH y otras veces contienen FSH, por lo que la liberación diferencial de LH y FSH debe depender de la interacción de varios factores (como GnRH, estradiol, inhibina). También la disparidad en las vidas medias de LH (20-30 min) y FSH (2-3 h) afecta a sus niveles circulantes.
De los esteroides ováricos, el 17b-estradiol es el inhibidor más potente de la secreción de gonadotropinas, y actúa tanto sobre el hipotálamo como sobre la hipófisis. La inhibina, producida por las células de la granulosa del ovario, suprimen específicamente la liberación de FSH. La extirpación de los ovarios provoca un rápido aumento de los niveles circulantes de LH y FSH; la administración de estradiol a mujeres hipoestrogénicas causa una rápida disminución de dichos niveles. Sin embargo, para que se produzca la ovulación, el estradiol debe ejercer un efecto positivo sobre la secreción de gonadotropinas. Los efectos de retroacción del estradiol parecen depender del tiempo y de la dosis. Al inicio de la fase folicular, en la hipófisis anterior hay cantidades relativamente pequeñas de LH y FSH disponibles para ser liberadas. Los niveles de estradiol (producido por el folículo seleccionado) aumentan, estimulando la síntesis de LH y FSH pero inhibiendo su secreción. En la mitad del ciclo, los niveles elevados de estradiol ejercen una retroacción positiva; éstos, con la GnRH y las cantidades bajas, pero en aumento de progesterona, inducen la oleada de LH. No se sabe si la liberación pulsátil de GnRH aumenta en la mitad del ciclo; el aumento en la mitad del ciclo podría resultar de un rápido incremento en el número de receptores (estimulados por estrógenos) presentes en las células productoras de gonadotropinas.
La menopausia, momento en el cual cesa la función cíclica del ovario manifestada por la menstruación, se trata en el capítulo 236.